|
|
Abstract
Objectives: UV irradiation may cause dermal ageing and carcinoma. The molecular action mechanism of such an effect and the methods for prevention are still lacking.
Materials and Methods: UVA (320-400 nm) plus UVB (290-320 nm) (hereafter denoted as UVAB) was used to induce skin damages in human fibroblast WS1 (hfWS1) cells. The parameters evaluated included the cell viability, expressions of relevant enzymes including MMP-1, MMP-2, catalase, and LDH; some biochemical indices such as elastin content and lipid peroxidation. Arbutin (AB) and ursolic acid (UA) were used to evaluate whether they could show any protective effect.
Results: UVAB inhibited the viability of hfWS1 cells, leading to a lowered elastin biosynthesis, enhanced release of LDH, and up-regulation of MMP-1, MMP-2 and catalase. Moreover it accelerated lipid peroxidation. In this regard, AB and UA behaved differently. On treatment with AB and/or UA, the cell viability was effectively protected by AB at dose <10 M and by UA at 1 M. In contrast, apparent cytotoxicity was shown by UA at 10 M. Although the extracellular elastin levels were recovered, yet
were insignificant. At a dose of 100 M, the lipid peroxidation was effectively suppressed by UA but not by AB. At 0.1 M, both AB and UA effectively suppressed the LDH release to the control level. Molecular action mechanism revealed both AB and UA at 1-2 M significantly down-regulated the expressions of catalase, MMP-2 but not MMP-1.
Conclusion: An appropriate combination of AB (100 M) and UA (5 M) would exhibit very strong protective effect on skin damages caused by UVAB. Introduction
Damages caused by UVB (290-320 nm) initially involve skin rash and ultimately severe skin carcinoma. The synergistic action of UVA (320-400 nm) and UVB occasionally would induce pronounced photoaging [1]. As well known, the action mechanism of UV-radiation is often initiated with production of tremendous reactive oxygen species (ROS) [2] including superoxide anion
(·O2-),
singlet oxygen
(1O2), hydrogen peroxide (H2O2), hydroperoxy free radical
(·HO2),
and hydroxyl free radicals
(·OH).
In view of radiation biology, a diversity of damaging effects could be induced by ROS, like DNA damage, lipid peroxidative degradation and protein denature, which could be summed up to trigger much default in signal transduction, gene transcription [3], and dermal aging. ROS directly destroy tissue collagens by inhibiting the tissue inhibitor of matrix-metalloproteinases (TIMPs) and the de novo synthesis of procollagens. ROS also can trigger the MAPK signal transduction pathway. ROS caused over-expression of matrix metalloproteinases (MMPs) and NFkB [4,5,6]. The latter in turn induced the secretion of MMP-8 (neutrophil collagenase) to accelerate degradation of dermal stratum. Pathologically, skin stromal substrate collagen and elastin are readily degradable by synergistic action of a diversity of proteases, like the interstitial collagenase (MMP-1), gelatinase-B (92 kDa MMP-9), gelatinase-A (72 kDa MMP-2), stromelysin-1 (MMP-3), membrane type-MMPs (MT-MMPs) and other un-classified MMPs [4,5,6]. Among which MMP-1, -2, and -3 are more readily inducible by UVA than UVB, the latter normally only triggers expression of MMP-1 and MMP-3. Pathobiochemically, photoaged skins are characteristics of skin wrinkling, loss of elasticity, skin roughness and thickening as well as abnormal dermal pigmentation. These pathophysiological indications usually are associated with many tissue- and molecular biological changes [7,8,9,10]. More progressively, epidermal thickening, connective tissue deformation and photocarcinogenesis were often seen in the late stage of UV damages [11,12]. Arbutin (I), popularly named hydroquinone- -D-glucopyranoside, has a chemical name of 2-(hydroxymethyl)-6-(4-hydroxyphenoxy) oxane-3,4,5-triol (Wikipedia, the free encyclopedia; Jump to: navigation, search).
I is commonly found in a diversity of food plants which has been used as a skin whitener and brightener since the past two decades. Chemically,
I is a glycosylated benzoquinone widely occurring in the bearberry plant genus Arctostaphylos. (Wikipedia, the free encyclopedia; "http://en.wikipedia.org/wiki/Arbutin"). When applied to skin, I readily infiltrates into skin without affecting any cell multiplication. And in epidermal layer it is slowly hydrolyzed to release hydroquinone, hence has been used as a very efficient skin lightener in cosmetics [13]. The reason is that the released hydroquinone in turn effectively suppresses melanin synthesis by inhibiting the enzyme tyrosinase [14]. Alternatively, ursolic acid
(II) is a pentacyclic triterpenoid having a chemical name of (3)-3-hydroxyurs-12-en-28-oic acid, which is commonly found in apples, blue berries, cranberries, peppermint, lavender, oregano, thyme, hawthorn and prunes.
II virtually is a cytostatic and an anti-proliferative [15,16], and more interestingly, it possesses a membrane stabilizing effect [17]. Both et al. [18] demonstrated that II increased both the ceramide and collagen contents in cultured normal human epidermal keratinocytes and dermal fibroblasts. It also accelerated lipid production, which in turn moisturized and strengthened the outer barrier for skin support (www.remergenskin.com). Pharmaceutically, the drawbacks of many antioxidant ingredients reside in high instability, easy oxidation and extremely large cytotoxicity. Proposing that
I and II could be bettter for use in this regard, we examined the UV-protective effects of
I and II respectively in human dermal fibroblast WS1 cells with an aim to define an appropriate dose in combined use of these two antioxidants.Materials and Methods
Cells Human dermal fibroblast WS1 cells were purchased from Food Industrial ATCC of Development Research Institute, Hsin-Chu, Taiwan.
Chemicals and Reagents Penicillin-streptomycin solution, non-essential amino acid solution (NEAA), 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), gelatin, Tris-HCl, sodium dodecyl sulfate (SDS), bromophenol blue, Triton-X, bis(N,N-methylene-bisacrylamine), acrylamide, N,N,N',N'-tetramethyl-ethylenediamine (TEMED), Coomassie Brilliant Blue R (CBR), butylated hydroxytoluene (BHT), thiobarbituric acid (TBA), ethylene dinitrilotetraacetic acid (EDNTA), NP-40, deoxycholate, hydrogen peroxide (H2O2), ammonium molybdate, LDH Assay Kit etc. were manufactured by Sigma (USA). Dulbecco's Modified Eagle Medium (DMEM) was purchased from Gibco Co. (USA). Dimethyl sulfoxide (DMSO) was provided by Riedel-De Haën Co. (Germany). Fetal bovine serum (FBS) was purchased form Biological Industries (Kibbutz Beit Haemek). Fastin Elastin Assay Kit was product of Biocolor Co. (Northern Ireland). Arbutin (AB), ursolic acid (UA), and (-)epigallocatechin gallate (EGCG) were obtained from Sigma-Aldrich.
Cell Cultivation Cells were plated onto 10 cm Petri dishes. DMEM incorporated with sodium bicarbonate, penicillin-streptomycin solution, FBS and NEAA was added. The incubation was conducted at 37oC under 5% CO2 atmosphere.
Induction by Arbutin and Ursolic Acid I and II were respectively dissolved in DMSO (5%) and diluted with 95% of PBS to a preliminary concentration of 10mM. The solution was further diluted with PBS to required concentrations of 0.10, 1.0 and 10μM in culture media. The cultivation was performed for 24 h at 37oC. After irradiated simultaneously with UVA (385nm, 1500
μw/cm2, 100mJ/cm2) plus UVB (302nm, 1500μw/cm2, 100mJ/cm2) (denoted as UVAB hereafter) for additional 24 h in the presence and the absence of
I and II, the cultures were examined for UV damages. Cell Viability In principle, when MTT is metabolized by mitochondrial dehydrogenase in living cells, formazan (the reduced form of MTT) would be produced, which is thus accumulated as insoluble purplish blue crystals in living cells
(Fig. 5). Briefly, according to Mosmann [19], human dermal fibroblast WS-1 cells were plated onto a 24 well plate at a density of 2×105 cell/well. After a 24 h of incubation, the cultures were
UVAB irradiated in the presence and the absence of I and II at concentrations as indicated. The cultures were incubated at 37
oC under 5% CO2 atmosphere. After cultivation for additional 48 h, the culture media were sucked off. To the culture 1 mL of MTT solution (0.5 mg/mL in DMEM) was added. The incubation was continued at 37oC under 5% CO2 atmosphere for additional 3.5 h. On having removed the culture media, 1 mL of DMSO was added. The mixture was agitated for 2 min. Aliquots of 100 L were respectively transferred into each well of a 96-well plate. The absorbance of each well was measured at 570 nm. The cell viability was calculated against the control.
Assay for MMP-1 Activity The assay method for MMP-1 was carried out using the Matrix Metalloproteinase 1 BioAssay ELISA kit (United States Biological) by following the manufacturer's instruction. Aliquots of 100 L sample solution were placed onto a 96-well plate. Each well of which had been previously placed with the human MMP-1 monoclonal antibodies. The incubation was continued for 2.5 h to facilitate the reaction. The supernatant was removed. After the cells were rinsed four times with rinsing solution, 100 L of secondary antibodies were added. The reaction was proceeded for additional 1 h. Cells were rinsed again four times. HRP-Streptavidin solution was added and the mixture was left to stand for 45 min to facilitate the reaction. The mixture was repeatedly rinsed for five times. To the culture 100
mL of TMB 3,3',5,'5-teramethylbenzide solution was added. After the mixture was left to stand for 30 min in the dark to avoid direct sunlight, the reaction was terminated by adding 50
mL of 2 M sulfuric acid. The absorbance was measured at 450 nm using an ELISA reader.
Assay for MMP-2 Activity The culture of WS-1 cells (2×105 cells/well) was UVAB irradiated in the presence and the absence of AB and UA. The culture media containing WS-1 cells were gently sucked out. To each 5 L of the culture, 5 L of tracer dye solution was added. The mixture was heated on water bath at 37
oC for 15min. The supernatant was decanted into a collagen-containing SDS-PAGE (7.5%), vortexed thoroughly and subjected to electrophoresis at 110 V for 60 min. The tracer dye was rinsed with 2.5% of Triton-X100. The plates were cultivated for 14 h in an incubation buffer (0.05 M Tris-HCl, 10 mM CaCl2, 50 mM NaCl, 0.05% Brij35) and then stained with CBR, and subsequently treated with the decolorizing solution (10% acetic acid plus 30% methanol). The decolorized areas on
the finished plates were scanned.
LDH Release In principle, lactic dehydrogenase (LDH) is a very stable intracellular enzyme in normal cells, which is readily released to extracellular plasma in case of mitochondrial damages. Hence the proportion of LDH released into the extracellular plasma can be used as a marker to evaluate the extent of cellular damages. In brief, WS-1 cells were plated onto a 24-well plate at a concentration of 2×104 cells/well. The cells were irradiated simultaneously with UVAB in the presence and the absence of I and II at concentrations indicated. The cells were incubated for 48 h. The supernatant media were decanted and centrifuged at 1500 rpm for 5 min. To an aliquot of 100
mL of the supernatant 50
mL of Kit Mixed Agent (In Vitro Toxicology Assay Kit, Sigma) were added. After incubation at ambient temperature for 30 min. Fifteen
mL of 1N HCl was added to terminate the reaction. On standing at ambient temperature for 10 min to facilitate the color reaction, the absorbance was measured at 490 nm.
Determination of Extracellular Elastin Elastin is normally secreted by skin fibroblast cells under physiological condition. On reaction with 5,10,15,20-tetraphenyl-21,23-porphine sulfonate (TPPS), a red precipitate is formed. Based on this principle to follow the instructions given by the manufacturers, the Fastin Elastin Assay Kit was used to evaluate the extracellular elastin content. Briefly, WS-1 cells were plated onto 24 well plate at a concentration of 2×104 cells/well. An aliquot of treated sample (100
mL) was added with 1 mL of Elastin Precipitating Reagent. The mixture was mixed thoroughly and refrigerated overnight at a temperature below 0oC. The reaction mixture was centrifuged (Hitachi, Himac CF 15R) at 10000×g for 20 min. The supernatant precipitating reagent was decanted. Two hundred L of 90 % saturated ammonium sulfate solution and 1 mL of Fastin Dye Reagent were added. The mixture was left to stand for 60 min to facilitate the reaction. The reaction mixture was centrifuged at 10000×g for 10 min. The supernatant was decanted. One mL of Fastin Dissociation Reagent was added and agitated vigorously to accelerate complete dissolution. An aliquot of 100
mL was measured and transferred into 96 well-plate. The absorbance was measured at 513 nm with ELISA reader. A reference
α-elastin was similarly treated to establish the calibration curve, against which the content of elastin was calculated.
Lipid Peroxidation in Cell Membrane On UVAB irradiation, the extent of lipid peroxidation can be readily assessed by determination of the intermediate lipid oxidative product, malondialdehyde (MDA) produced in parallel. Briefly, WS-1 cells were plated onto a 6-well plate at a concentration of 1×106 cells/well. I and II were added, respectively, at concentrations as indicated. After incubated for 24 h, the cultures were irradiated with UVAB. The cells (0.5 mL) were harvested and treated according to Tsujimura (1990) [20], i.e. 12.5mL of 4% BHT, 125 L of 0.3% of SDS, 0.5mL of 0.5N HCL and 0.25mL of 0.8% TBA were successively added. After thoroughly agitated, the mixture was heated in water bath (100oC) for 45 min and then cooled to room temperature. To the final reaction mixture 0.75 mL of 1-butanol was added. The mixture was left to stand for 20 min. The supernatant was separated, and the absorbance was measured at 532 nm.
Determination of the Intracellular Lipoxygenase Activity Cells were harvested by centrifugation at 3000 rpm for 5 min. The supernatant was decanted. To the residue RIPA Buffer was added. The mixture was left to stand for 30 min at 4
oC and then centrifuged at 12000 rpm for 10 min. The supernatant was separated for further use. Fifty
mL of the sample solution was mixed with 50mL of substrate H2O2 solution (6.5
mmol). The mixture was mixed thoroughly to facilitate the reaction at ambient temperature for 60s. To the final reaction mixture 100mL of ammonium molybdate solution was added. The absorbance was read at 405 nm using an ELISA reader.
Results
Cell Viability Protective Effect of Arbutin and Ursolic Acid Against UV-Irradiation When exposed to UVAB, cell viability was reduced by 40 % if without treatment
(Fig. 1). At 0.1
μΜ, arbutin and ursolic acid rescued cell viability by 22 % and 55 %, respectively
(Fig. 1). Both arbutin and ursolic acid showed very satisfactory UV-protective effect comparing to EGCG at doses below
10μM. Differently, ursolic acid at dose
³10μΜ
apparently exhibited strong cytotoxic effect, which was totally not found for arbutin
(Fig. 1). Obviously, arbutin was much safer than ursolic acid regarding a wider dosage spectrum for application. Thus, cell viability can be protected by arbutin at doses <10 M, while by ursolic acid at doses approximately within 0.5 M.
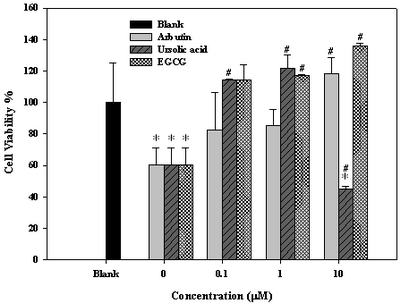 | Fig 1:
Effects of arbutin, ursolic acid, and EGCG on viability of human
dermal fibroblast WS-1 cells on exposure to UVAB irradiation.a
aData expressed in mean±S.D. from
triplicate experiments. *p<0.05 compared with blank, #p<0.01
compared with control.
UVAB irradiation =UVA (385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Concentration at 0mM: cells treated
with UVAB irradiation in the absence of AB, UA and EGCG.
Concentration at 0.1-10mM: cells treated with UVAB irradiation in
the presence of 0.1 to 10mM of AB, UA, and EGCG, respectively. EGCG
was used as the positive control |
|
MMP-2 Was Down-regulated by Arbutin and Ursolic Acid On exposure to UVAB, MMP-2 in fibroblast cells was significantly up-regulated
(Fig. 2), which was significantly ameliorated by arbutin and ursolic acid at doses 1.0 M, results being comparable to that of EGCG
(Fig. 2).
 |
Fig 2:
Effect of arbutin, ursolic acid, and EGCG on expression of MMP-2
in human fibroblast WS-1 cells after irradiation with UVABa
a UVAB irradiation =UVA (385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Concentration at 0mM: cells treated
with UVAB irradiation in the absence of AB, UA and EGCG.
Concentration at 1mM: cells treated with UVAB irradiation in the
presence of 1mM of AB, UA and EGCG, respectively. EGCG was used as
the positive control. |
|
UVAB Activated MMP-1 Which Was Further Enhanced by Arbutin and Ursolic Acid The normal physiological level of MMP-1 was around 0.0092pg/mL, when induced by UVAB the level was activated to reach 0.0100 pg/mL, which was seen to be further up-regulated by arbutin and ursolic acid at a dose of 1 M to 0.0121 pg/mL and 0.0126 pg/mL, respectively
(Fig. 3). In this regard, arbutin revealed activity comparable activity to that of EGCG
(Fig. 3).
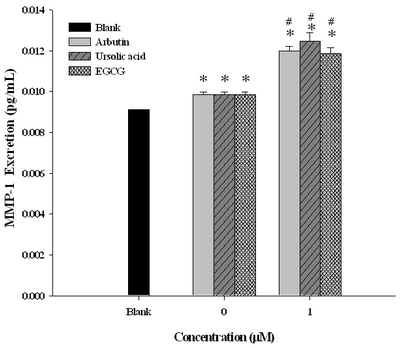 | Fig
3:
Effects of arbutin, ursolic acid, and EGCG on expression of MMP-1
collagenase in human dermal fibroblast WS-1 cells after irradiated
with UVAB a
a Data expressed in mean±S.D. from
triplicate experiments. *p<0.01 compared with blank, #p<0.05
compared with the control.
UVAB irradiation =UVA (385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Concentration at 0mM: cells treated
with UVAB irradiation in the absence of AB, UA and EGCG.
Concentration at 1mM: cells treated with UVAB irradiation in the
presence of 1mM of AB, UA and EGCG, respectively. EGCG was used as
the positive control. |
|
LDH Release Was Efficiently Inhibited by Arbutin and Ursolic Acid In normal cells the extracellular LDH level was maintained approximately at a steady level of 34.68%, while exposure to UVAB triggered the LDH release to a level of 56.74%. At a dose of 0.01μΜ
, arbutin and ursolic acid respectively suppressed the LDH release to 35.28 and 40.09%, comparing to 42.40% by EGCG (Table 1). When the dose was increased to 0.1μΜ, ursolic acid was seen to further effectively inhibit the LDH release to 33.13% and similarly to 37.00% by EGCG, however, unlikely with arbutin
(Table 1). Interestingly, both arbutin and ursolic acid showed slightly better results than EGCG (Table 1).
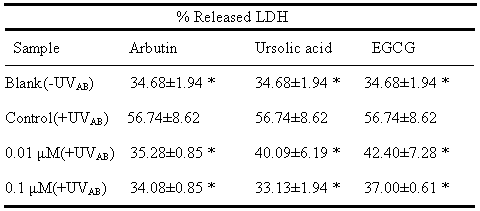 |
a Data
expressed in mean±S.D. from three experiments. *p<0.05 was
the significant level compared with control.
UVAB irradiation =UVA
(385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Control: cells treated with UVAB
irradiation in the absence of AB, UA and EGCG. Concentration at 0.01
to 0.10mM:
cells treated with UVAB irradiation in the presence of
0.01-1.0M of AB, UA, and EGCG, respectively. EGCG was used as the
positive control. |
|
Arbutin and Ursolic Acid Restored Elastin Biosynthesis Post UV Irradiation As well recognized, an alternate damaging effect of UVAB actually targets on elastin biosynthesis. The content of elastin was seen suppressed by 10 % (from 47mg for blank to 37 % for the untreated) on exposure to UVAB
(Fig. 4). In this respect, arbutin and ursolic acid revealed to be far less effective
(Fig. 4).
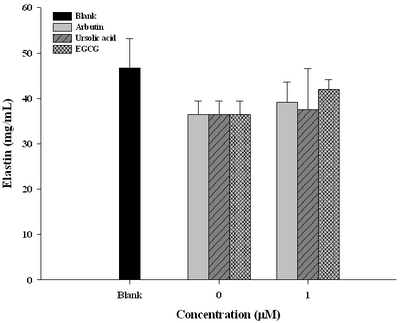 | Fig
4:
Restoration of elastin secretion in human fibroblast WS-1 cells
by arbutin, ursolic acid, and EGCG after UVABa irradiation a
a UVAB irradiation =UVA (385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Concentration at 0mM: cells treated
with UVAB irradiation in the absence of AB, UA and EGCG.
Concentration at 1mM: cells treated with UVAB irradiation in the
presence of 1mM of AB, UA, and EGCG, respectively. EGCG was used as
the positive control.
|
|
Arbutin and Ursolic Acid Suppressed Lipid Peroxidation In Cell Membrane Taking the control sample (irradiated, but without antioxidant treatment) as 100%. Arbutin alone at dosages either 50 M or 100 M was totally ineffective to inhibit lipid peroxidation. In contrast, ursolic acid and EGCG revealed significant suppressions, reaching 34.5% and 51.7% at 50 M, and 51.7% and 58.6% at 100 M, respectively
(Fig. 5). Such a deviation in anti-lipid peroxidative activity could be ascribed to solubility difference between arbutin and ursolic acid. Arbutin is more water soluble due to the carriage of glucose moiety in its structure, and this is the reason it failed a lot in assessing the LDL fraction.
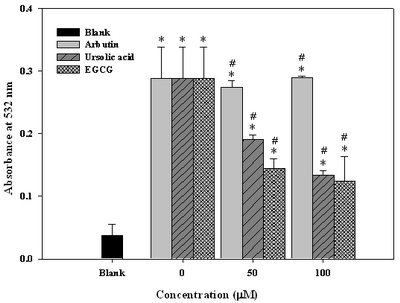 | Fig
5:
Effect of arbutin, ursolic acid and EGCG on lipid
peroxidation in human fibroblast WS-1 cells after irradiated with
UVAB a
a Data expressed in mean±standard
deviation from triplicate experiments. *p<0.05 compared with
control, #p<0.01 compared with the blank.
UVAB irradiation =UVA (385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Concentration at 0mM: cells treated
with UVAB irradiation in the absence of AB, UA and EGCG.
Concentration at 50-100mM: cells treated with UVAB irradiation in
the presence of 50-100mM of AB, UA, and EGCG, respectively. EGCG was
used as the positive control. |
|
Arbutin and Ursolic Acid Inhibited Catalase Activity in Cell Membrane UVAB triggered up-regulation of catalase activity in membrane of WS-1 fibroblast cells. reaching a two-fold increase in activity (Fig. 6), which was effectively inhibited by arbutin and uesolic acid at 1
μΜ by 70 % and 40 %, respectively
(Fig. 6).
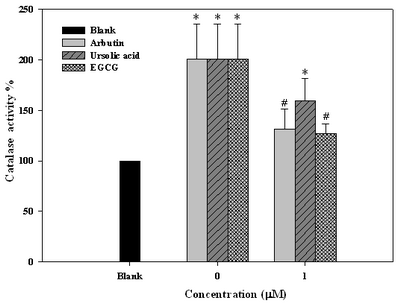 | Fig
6:
Effect of arbutin, ursolic acid and EGCG on catalase expression
in human fibroblast WS-1 cells after irradiation with UVABa
aData expressed in mean±S.D. from
triplicate experiments. *0.1>p>0.05 compared with the blank, #p<0.01
compared with the control.
UVAB irradiation =UVA (385nm, 1500 μw/cm2, 100 mJ/cm2) plus UVB
(302nm, 1500 μw/cm2, 100mJ/cm2).
Blank: without any treatment. Concentration at 0mM: cells treated
with UVAB irradiation in the absence of AB and UA. Concentration at
1.0mM: cells treated with UVAB irradiation in the presence of 1.0M
of AB, UA, and EGCG, respectively. EGCG was used as the positive
control. |
|
Discussion
Recently, the carcinogenesis of UV-irradiation has attracted many researchers. As huge amount of reactive oxygen species (ROS) can be generated on exposure to UV-irradiation to injure epidemic cells,
such oxidative damages on skin always require a relatively long-term exposure to ROS, which in turn would eventually lead to serial physiological hazards including the suppressed cell viability [21], intracellular lipid perooxidation, and formation of huge amount of malondialdehyde, a common oxidative intermediate of a number of lipids [22]. Steinbrenner et al. [23]
demonstrated that UVAB up-regulates expression of MMP's through induction of a cascade of signal transduction including the transcription factor AP-1, which in turn induces the expression of MMP's and accelerates the in vivo degradation of elastin and collagens, eventually leading to dermal
ageing [3,23]. In this present study, the decrease in cell viability caused by UVAB irradiation was shown effectively restored by treatment with arbutin at dose 10 M and ursolic acid at dose <5 M. Alternatively, in agreement with Lee et al. [24], the cytotoxicity of ursolic aicd at dosages 10 M obviously would severely limit the application of ursolic aicd (Fig. 1). Arbutin was slightly less effective in MMP-2 inhibition, comparing to ursolic acid and EGCG (Fig. 2). On the contrary, MMP-1 was significantly over-expressed by both arbutin and ursolic acid
(Fig. 3). Similar results had been confirmed by Moon et al. [25]. Alternatively, the up-regulation of MMP-1 could also be attributed to the cell viability protective effects of arbutin and ursolic acid. Cytophysiologically, the up-regulation of MMP-1 could mean more survival cells. Otherwise, the amount of MMP-1 determined by ELISA kit straight forwardly only represents the total protein, which actually involves both parts: the activated MMP-1 and the non-activated pro-MMP-1. Because the total protein content as presently determined only roughly reflects the activity of MMP-1. Hence a more accurate criticizing may require a sharper and more sensitive kit other than zymography. In addition, UV-irradiation can cause cell membrane damages with enlarged membrane porosities, resulting in enhanced permeability or flux, hence larger amount of LDH was able to be transiently released from the intracellular to the extracellular compartment. Accordingly, the amount of LDH released can serve a direct index of the extent of cell damages. As seen, arbutin and ursolic acid efficiently retarded LDH release in a dose-responsive fashion, giving strong support of the protective activity of these two nutraceuticl compounds
(Table 1). Alternatively, although arbutin and ursolic aicd seemed to have been able to restore elastin content in fibroblast cells
(Fig. 4), yet still insignificantly. Uniquely, ursolic acid was effective to suppress lipid peroxidation, however arbutin was unlikely
(Fig. 5). As for antioxidative activity, consistent with Milton [26], the up-regulation of calatase in fibroblast cells was significantly suppressed by arbutin and ursolic acid
(Fig. 6). Practically, the skin does not exist in isolation from the rest of the body. Hence, skin rejuvenation has a much better chance of success as a part of a comprehensive anti-aging strategy. Even if the skin could be rejuvenated independently, what good is a 20-year old looking skin in the body that feels eighty. Besides, the anti-aging measures for the body benefit the skin too. Hence, the protection of skin is suggested to cover the whole body aging concept. Conclusively, arbutin and ursolic acid are good UV-protectors, their molecular action mechanism can be ascribed to i) protecting the cell viability; ii) down-regulating the MMP-2; iii) up-regulating the MMP-1; iv) elevating the extracellular elastin levels. v) suppressing the LDH release; vi) inhibiting the lipid peroxidation; and vii) down-regulating the catalase. And finally, a combination of arbutin ( 100 M) and ursolic acid ( 5 M) may attain an optimal concentration to exhibit the most potent UV-protective effect in delaying the skin photoaging.
Conflict of Interest Statement: All authors declare that there are no any actual or potential conflict of interest including any financial, personal or other relationships with other people or organizations within that could inappropriately influence (bias) our work. References
1.
Diffey BL. Human exposure to ultraviolet radiation. In: HawkJLM, ed.
Photodermatology. London: Arnold, 1999.
2.
Herrling Th, Jung K, Fuchs J. Measurements of UV-generated free
radicals/reactive oxygen species (ROS) in skin. Spectrochimica Acta Part A
2006; 63: 840- 5.
3.
Rittié L, Fisher GJ. UV-light-induced signal cascades and skin aging. Ageing
Res Rev 2002; 1: 705- 20.
4.
Chambers AF, Matrisian LM, Changing views of the role of matrix
metalloproteinases in metastasis. J Natl Cancer Inst 1997; 89: 1260- 70.
5.
Zhou LZ, Johnson AP, Rando TA. NF-B and ap-1 mediate transcriptional
responses to oxidative stress in skeletal muscle cells. Free Rad Biol Med
2001; 31: 1405- 16.
6.
Onoue S, Kobayashib T, Takemoto Y, Sasaki I, Shinkai H. Induction of matrix
metalloproteinase-9 secretion from human keratinocytes in culture by
ultraviolet B irradiation. J Dermatol Sci 2003; 33: 105- 11.
7.
Gilchrest BA. Skin aging and photoaging: an overview. J Am Acad Dermatol
1989; 21: 610- 3.
8.
Fisher L, Bernstein EF, Chen YQ, Kopp JB, Brown DB, Hahn PJ, Robey FA,
Lakkakorpi J, Uitto J. Long-term sun exposure alters the collagen of the
papillary dermis. Comparison of sun-protected and photoaged skin by northern
analysis, immunohistochemical staining, and confocal laser scanning
microscopy. J Am Acad Dermatol 1990; 34: 209- 18.
9.
Varani J, Warner RL, Gharaee-Kermani M, Phan SH, Kang S, Chung JH, Wang ZQ,
Datta SC, Fisher G.J, Voorhees JJ. Vitamin A antagonizes decreased cell
growth and elevated collagen-degrading matrix metalloproteinases and
stimulates collagen accumulation in naturally aged human skin. J Invest
Dermatol 2000; 114: 480- 6.
10.
Wlaschek M, Tantcheva-Poo´r I, Naderi L, Ma W, Schneider LA. Razi-Wolf, Z.,
Schu¨ller, J., Scharffetter-Kochanek, K. Solar UV irradiation and dermal
photoaging. J Photochem Photobiol B: Biology 2001; 63: 41- 51.
11.
Beissert S, Granstein RD. UV-induced cutaneous photobiology. Crit. Rev.
Biochem. Mol. Biol 1996; 31: 381- 404.
12.
Scharffetter-Kochanek K, Wlaschek M, Brenneisen P, Schauen M, Blaudschun R,
Wenk J. UV-induced reactive oxygen species in photocarcinogenesis and
photoaging. Biol Chem 1997; 378: 1247- 57.
13.
Devillers J, Boule P, Vasseur P, Prevot P., Steiman, R, Seigle Murandi F,
Benoit-Guyod JL, Nendza M, Grioni C, Chambon P. Environmental and health
risks of hydroquinone. Ecotoxicol Environ Safe 1990; 19: 327- 54.
14.
Sugimoto K, Nishimura T, Nomura K, Sugimoto K, Kuriki T, Inhibitory effects
of alpha-arbutin on melanin synthesis in cultured human melanoma cells and a
three-dimensional human skin model. Biol Pharm Bull 2004; 27: 510- 4.
15.
Cha HJ, Park MT, Chung, HY. Ursolic acid-induced down-regulation of MMP-9
gene is mediated through the nuclear translocation of glucocorticoid
receptor in HT1080 human fibrosarcoma cells. Oncogene 1998; 6: 771- 8.
16.
Es-Saady D, Simon A, Jayat-Vignoles C, et al. MCF-7 cell cycle arrested at
G1 through ursolic acid, and increased reduction of tetrazolium salts.
Anticancer Res 1996; 16: 481- 6.
17.
Han, S.K., Yi, K.O. Park, S.J. Oleanolic and ursolic acid stabilize
liposomal membranes. Lipids 1997; 32: 769- 73.
18.
Both, D.M., Goodtzova, K., Yarosh, D.B. Brown D A. Liposome-encapsulated
ursolic acid increases ceramides and collagen in human skin cells. Arch.
Dermatol Res 2004; 293: 569- 75.
19.
Mosmann T. Rapid colorimetric assay for cellular growth and survival:
application to proliferation and cytotoxicity assays. J Immunol Methods
1983; 65: 55-63.
20.
Tsujimura T, Maher VM, Godwin AR, Liskay RM, McCormick JJ. Frequency of
intrachromosomal homologous recombination induced by UV radiation in
normally repairing and excision repair-deficient human cells. Proc Natl Acad
Sci U S A 1990; 87: 1566- 70.
21.
Halliwell B. Reactive oxygen species in pathology with special reference to
the skin. In: Fuchs J, Packer L,(eds.): Oxidative Stress in Dermatology,
Dekker Publishing, New York, 1993.
22.
Stratton SP, Liebler DC. Determination of singlet oxygenspecific versus
radical-mediated lipid peroxidation in photosensitized oxidation of lipid
bilayers: effect of beta-carotene and alpha-tocopherol. Biochemistry 1997;
36: 12911- 20.
23.
Steinbrenner H, Ramos MC, Stuhlmann D, Sies H, Brenneisen P. UVA-mediated
downregulation of MMP-2 and MMP-9 in human epidermal keratinocytes. Biochem
Biophys Res Commun 2003; 308: 486- 91.
24.
Lee YS, Jin DQ, Beak SM, Lee ES, Kim JA. Inhibition of ultraviolet A
modulated signaling pathways by asiatic acid and ursolic acid in HaCaT human
keratinocytes. Eur J Pharmacol 2003; 476: 173- 8.
25.
Moon HI, Lee J, Eun HC, Kim KH, Chung JH. The effect of tiarellic acid on
the expressions of matrix metalloproteinase-1 and type 1 procollagen in
ultraviolet irradiated cultured human skin fibroblasts. J Ethnopharmacol
2005; 98: 185- 9.
26.
Milton NG.N. Inhibition of catalase activity with 3-amino-triazole enhances
the cytotoxicity of the alzheimer's amyliod-β peptide. Neuro Toxicology
2001; 22: 767-74© 2009 Egyptian Dermatology Online Journal |

